Early Blight of Potato
(PP1892, Reviewed Nov. 2025)Early blight is caused by the fungal pathogen Alternaria solani. The disease affects leaves, stems, and tubers, and can reduce tuber yield and size. Reductions in tuber quality due to early blight compromise storage longevity and marketability of fresh-market and processing crops.
Symptoms
Early blight foliar lesions can be diagnosed in the field by the characteristic dark concentric rings alternating with bands of light-tan tissue, giving them a distinctive target spot appearance (Figure 1a). Initial symptoms of early blight appear as small, circular or irregular dark brown to black spots, typically starting on the older (lower) leaves (Figure 1b). These spots enlarge up to 3/8 inch in diameter and gradually may become angular in shape, often restricted by leaf veins as they expand.


Initial early blight lesions may be confused with brown spot (caused by small-spore Alternaria species) (Figure 2a) or black dot (caused by Colletotrichum coccodes) lesions (Figure 2b). However, brown spot and black dot lesions do not develop dark concentric rings characteristic of early blight infection. Brown spot lesions rarely become larger than 3-4mm and are typically round. First early blight lesions appear about two to three days after infection, with sporulation at the lesion margin occurring three to five days later.


Multiple lesions on the same leaf may coalesce, or grow together, to form what appears to be one large lesion (Figure 3a), potentially resembling grey mold caused by Botrytis cinerea (Figure 3b). Chlorosis (yellowing of plant tissue) may be visible around clusters of infections by the early blight pathogen (Figure 3a). Elongated, brown to black lesions may develop on the stems and petioles of infected plants (Figure 4).



Later in the growing season, as infections become more severe, lower leaves may drop, and numerous lesions may appear on the upper leaves (Figure 5).

Premature leaf senescence, reduced yield and low tuber dry matter content are likely when plants suffer from severe foliar infection during the tuber bulking stage.
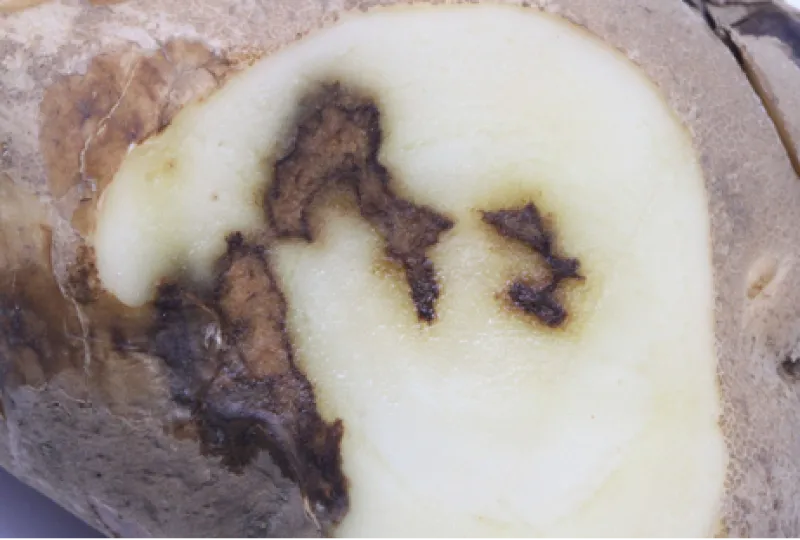
Early blight tuber symptoms appear as dark and sunken lesions on the surface (Figure 6). Tuber lesions may be circular or irregular in shape and surrounded by a raised dark brown border (Figure 7).


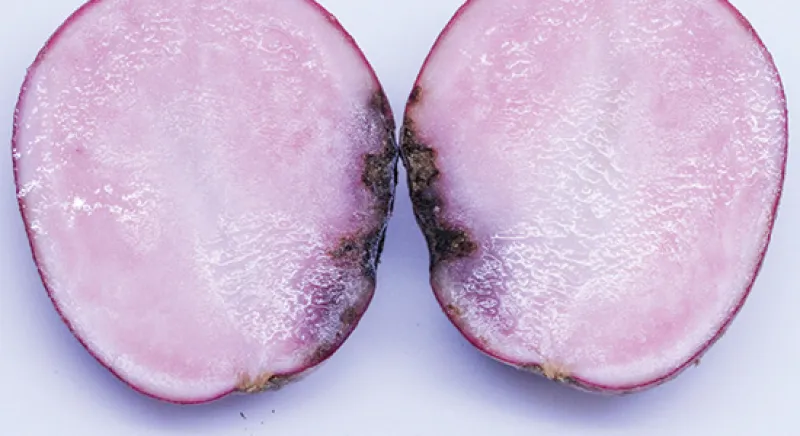
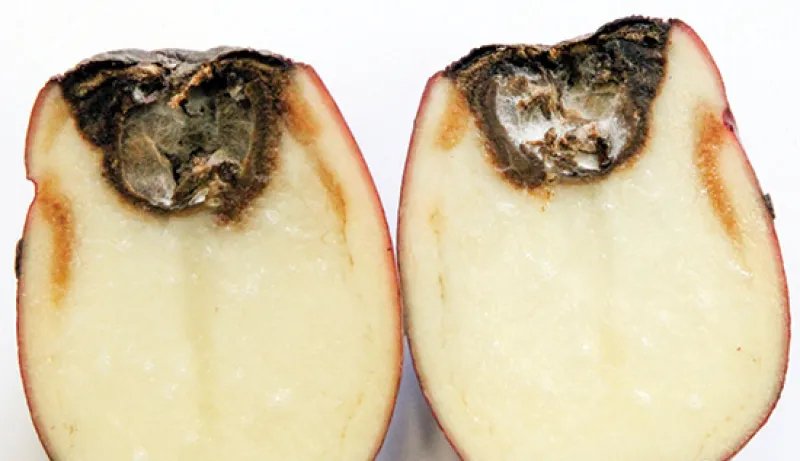
The underlying tuber tissue is dry and dark brown with a corky texture (Figure 8). Tuber symptoms of early blight may manifest only after months of storage (Figure 9) and can be confused with Fusarium dry rot (Figure 10).
Disease Cycle
A. solani inoculum for primary infections of potato foliage most commonly originates from spores (conidia) on plant debris from previous seasons, typically in neighboring fields. Other less common sources of pathogen inoculum include infected seed tubers, volunteer potatoes or alternative hosts such as tomatoes and hairy nightshade. Overwintering spores that serve as the initial inoculum move within and between fields carried by air currents, windblown soil and plant debris, splashing rain and irrigation water. Spores can survive freezing temperatures on or just below the soil surface.
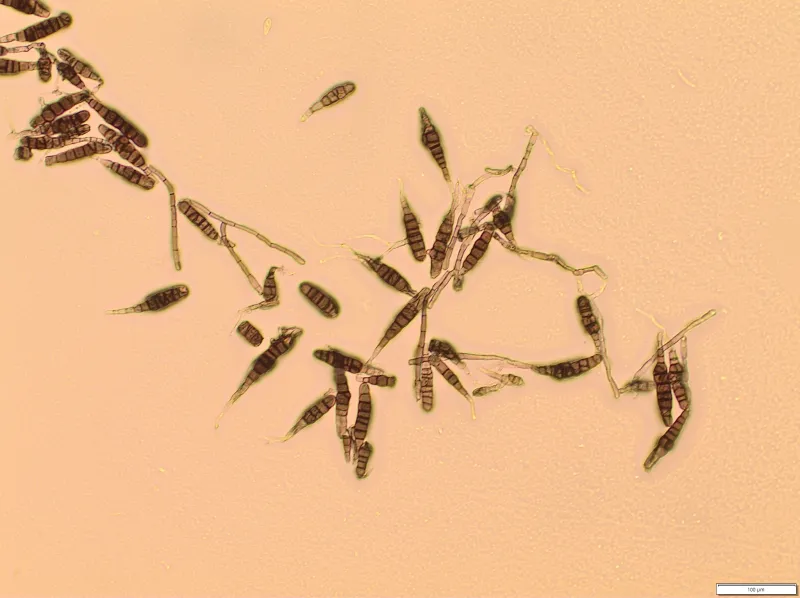
A. solani spores formed on infected foliage are dislodged during conducive environmental conditions (Figure 11). Alternating wet and dry periods are most favorable for spore formation, dispersal and the spread of the fungus to healthy tissue. Multiple sporulation and infection cycles occur within a single season, leading to an exponential increase in foliar disease as the growing season progresses. The infection rate is generally low in the early season, and increases after flowering and is most rapid during tuber bulking (Figure 12).


The optimum temperature for spore germination and infection by A. solani is 68 to 86 F, although it can occur between 41 and 95 F. Sporulation and disease development occur between 50 and 95 F, with the optimum temperature of 72 to 81 F. Because of the favorable wet/dry cycle, peak spore dissemination occurs during midmorning and declines throughout the afternoon into night.
High humidity or free moisture and favorable temperatures are required for spore germination and penetration into susceptible plants. Penetration can occur directly through host epidermal cells or via natural openings like stomata or wounds. The pathogen favors older, senescing leaf tissue. Plants stressed by injury, nutrient deficiency and insect feeding are highly vulnerable to infection.
During harvest, tubers often come into contact with A. solani spores that have accumulated on the soil surface during the growing season or were dislodged from desiccated vines. Spores germinate and penetrate tubers through lenticels and wounds caused by mechanical injury (Figure 13).

Tuber infection is most common in potato cultivars highly susceptible to skinning, such as red- and white-skinned cultivars. Infection does not spread during storage and, unlike late blight tuber lesions, early blight tuber lesions typically do not serve as infection courts for other decay organisms.